




The sometimes-confusing world of taxonomy helps botanists to understand the phylogenetic journey of a plant. In this article, we explore the phylogeny (evolutionary relationships) of cannabis and its evolution into the Cannabaceae family. We talk about cannabis ancestors and close relatives, and other members of the Cannabaceae family.
- Cannabis within the cladistic system
- The complex and confusing world of plant taxonomy
- Characteristics of the various taxa
- The Rosidae superorder and the Rosaceae family
- The relatively recent Urticalean rosids
- The confusing classification of the Cannabaceae family
- Other noteworthy Cannabaceae plants
- Are cannabinoids unique to Cannabis?
The task of categorizing and cataloguing the billions of plant species that have existed on our planet since the dawn of life is a huge and vastly complex one. Prior to the development of genetic testing, classification on the basis of perceived similarity was the only way to assign a plant to its correct taxon.
Now that we have developed the study of phylogenetics (evolutionary relationships between groups of organisms), we have some basis for classifying plants according to their genetic similarities — but that does not provide a problem-free approach to such a complex task.
Cannabis within the cladistic system
However, we can at least begin to classify plants according to a cladistic system. The cladistic system relies on the presence of a single common ancestor. In this way, every clade (branch) of the phylogenetic tree contains the descendants of a single common ancestor. When many plants share a common ancestor, they are said to be monophyletic.
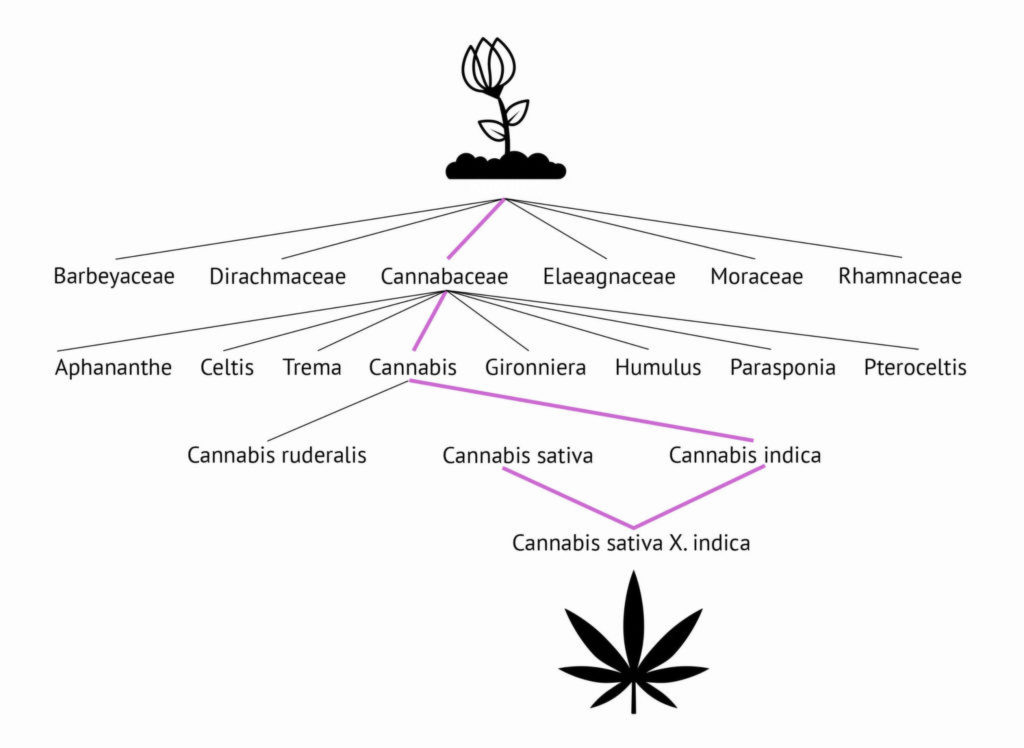
According to the Integrated Taxonomic Information System (ITIS), the species Cannabis sativa L. belongs to the genus Cannabis, which belongs to the Cannabaceae family. The Cannabaceae family, along with three other families, make up the (informal) suborder, Urticalean rosids, of the order Rosales. Along with the Urticalean rosids, another five families belong to the Rosales order; these include Rosaceae (rose) and Rhamnaceae (buckthorn).
Related post

The complex and confusing world of plant taxonomy
The Rosales order belongs to the informal subclass (or superorder) Rosanae (or Rosidae), a huge clade that comprises almost one quarter of all flowering plants. These ‘rosids’ belong to the Magnoliopsida or Eudicot class, which is a member of the Angiospermae infradivision. In turn, these belong to the subdivision of Spermatophytina, of the division of Tracheophyta, of the infrakingdom Streptophyta or Embryophyta (land plants), of the Viridiplantae (green plant) subkingdom.
The Viridiplantae phylum consists of green algae as well as all land plants. Along with the phyla of Rhodophyta (red algae) and Glaucophytes (freshwater microalgae), it makes up the Archaeplastida kingdom, which is also known as Plantae sensu lato — meaning “plants in the broader sense”.
The issue of whether Archaeplastida is in fact a higher clade comprised of the kingdoms of Viridiplantae etc. is somewhat controversial.
Characteristics of the various taxa
Archaeplastida all feature chloroplasts, which undergo photosynthesis to produce energy; furthermore, the chloroplast cells of archaeplastids share common structures, typically surrounded by just two membranes.
Viridiplantae differ from other phyla in the Archaeplastida clade in that they produce both chlorophyll a and b, giving them their usual green colour; Rhodophyta and Glaucophytes produce just chlorophyll a.
Embryophyta comprise all land plants, and are believed to have diverged from green algae around 540 million years ago (Ma).
Tracheophyta, or vascular plants, contain fibrous tissue to conduct water and minerals, and include mosses and ferns as well as flowering plants. Spermatophytina are vascular plants that produce seeds, and are believed to have diverged from non-seed-bearing plants around 319 Ma.
Angiospermae are classified thus due to their shared characteristic of bearing seeds enclosed in protective structures; they all bear flowers, which later produce the seed-containing fruit, and appeared on the fossil record around 192 Ma. Eudicots are angiosperms that express two embryonic leaves, as opposed to monocots that produce just one. Eudicots also exhibit a characteristic three-grooved pattern on the pollen grain.
Related post

The Rosidae superorder and the Rosaceae family
The Rosidae are believed to have appeared 108-121 Ma, and are usually characterised by their showy flowers—the Urticalean rosids are notable exceptions to this rule. Research has shown that the more recent evolutionary divergences have tended towards non-showiness, as wind-pollination became more prevalent. The Rosales order comprises a huge variety of evergreen and deciduous trees, shrubs, vines and herbs: for example, the Rosaceae family contains an incredible variety of important species including peach, apple, strawberry, almond, and of course rose.

The relatively recent Urticalean rosids
Thought to be relatively recent offshoots of the phylogenetic tree, the Urticalean rosids are characterised by their often non-showy and mostly unisexual flowers (although elm flowers are usually bisexual). As well as the Cannabaceae, the Urticales include the Urticaceae (nettle), Ulmaceae (elm), and Moraceae (mulberry or fig) families. Male flowers are inconspicuous and contain no petals, only sepals; female ovaries contain a single ovule, and produce a single seed.
Urticalean rosids exhibit huge variation in morphology and biogeography, and have developed some striking adaptations. Plants are usually wind-pollinated, although several Moraceae family members are insect-pollinated. Several species within the nettle family have the unique ability to explosively disperse pollen into the wind. Fruits may be hard, dry achenes, as in cannabis; fleshy drupes, as with hackberry; fleshy synconiums, unique to figs; or compound clusters as found in mulberry. Interestingly, several mulberry species are thought to contain mild hallucinogenic properties, and high-quality fibre can be obtained from the bark.
Related post

The confusing classification of the Cannabaceae family
The 170 or so members of the Cannabaceae family have few characteristics that are distinct from the Urticales as a whole. DNA evidence has shown that all are related, and certainly humulus (hops) and cannabis share obvious similarities with regard to floral structure. Both contain terpenoids, accounting for their similar fragrance, but humulus is a vine (climbing plant) and cannabis is herbaceous. The Celtis (hackberry) genus is relatively dissimilar: Celtis species are usually tall deciduous trees, with simple leaves, unlike the compound palmate leaves found in humulus and cannabis. Celtis trees are usually not dioecious and although flowers are usually unisexual, they are found on the same plant.

Despite the many differences, the male flowers of many Celtis species bear a striking resemblance to those of cannabis. Other similarities between members of the Cannabaceae family include stipule-bearing leaves (in cannabis, the stipules are the two tiny shoots at the base of each leaf), and cystoliths, or enlarged leaf cells containing crystals of calcium carbonate.
Other noteworthy Cannabaceae plants
Although it is commonly believed that Cannabis and Humulus are the only two Cannabaceae genera with any commercial value, there are several other species that are of great value in the regions in which they are indigenous.
Various Celtis species are cultivated either for timber or as ornamentals, such as the protected South African tree, C. Africana. It’s attractive, fast-growing and frost- and drought-resistant. Its abundant flowers and small, sweet fruits also attract a variety of birds and insects to the garden.
C. sinensis from East Asia and C. australis from southern Europe are also widely cultivated for these purposes.
Trema orientalis, a common species distributed throughout the tropical and warm temperate regions of Africa, Asia and Oceania, has abundant uses. Many indigenous populations refer to it as the “charcoal tree”, as one of its uses is for charcoal production. Its wood burns quickly and easily, and is also useful for making paper and pulp, due to the high tensile strength of the fibres.
The fibres of the bark are used to make rope or twine, and the sap exuded can be used to waterproof fishing lines. It is also used as animal fodder. Interestingly, T. orientalis has various traditional medicinal uses, many of which are quite similar to those of Cannabis—it is used to treat dysentery, inflammation, sore throats, and asthma, among other things, and has shown promising results in preliminary trials as a blood-sugar stabiliser in diabetes.
Aphananthe aspera, an East Asian species also known as the Muku tree, is a highly-prized ornamental traditionally used in Chinese classical gardens. A. aspera is also useful as a source of fibre and wood, and its leaves are used as fine sandpaper for polishing wooden and similar objects. A particularly fine, seven-trunked A. aspera is listed as one of South Korea’s natural monuments; this sixteen-metre specimen is believed to be around three hundred years old.

Are cannabinoids unique to Cannabis?
The Cannabis genus is apparently unique within the Cannabaceae family for containing cannabinoids. There is evidence to suggest that cannabimimetics – cannabinoid-like compounds – are found in other plant species, particularly echinacea, which is a member of the Asterid subclass, and must therefore have evolved separately to cannabis for millions of years. This may imply that a proto-cannabinoid system existed in a common ancestor of both plants, prior to the divergence of the Rosid and Asterid clades around 126 Ma.
However, as far as current knowledge goes, no other plant species produces THC or CBD. These cannabinoids are specific to the cannabis plant. There are companies who claim to have produced CBD from sources such as limonene from citrus fruit, and the bark of evergreens from Southeast Asia, but as yet, no independent peer-reviewed science has emerged to verify these claims.
Given the great improbability of a new plant species containing a fully functional phytocannabinoid system suddenly springing into existence, gradual evolution of complex cannabinoid systems from more rudimentary lineages seems to be the only plausible explanation. Traces of early Cannabaceae species have been found in Cretaceous fossils dating from up to 93.5 Ma, but there is no concrete evidence as to when the Cannabis genus diverged.
The fact that no close living relatives of cannabis show evidence of a cannabinoid system is surprising, but as research continues, more cannabinoid-containing plants may be discovered. This may necessitate further reshuffling of the complex and confusing system of plant classification.

I’m impressed. Will read this several times. Plus share. Big respect!
Rich people is best in the world and you want rich and make money more earn life enjoy.
Thank you very much! Very useful for school paper 🙂
I would say that cannabis is from the Urticales order. I would do a bit more research.
I am reading up on botanical nomenclature for college. The subject matter can be a little uninspiring! I thought I would try reading up on some of my favourite varieties and their taxonomy so I might relate and remember more. This article was the most informative yet. Bookmarked. +rep to the author.
Hello Pistil Pete,
Thank you for your comment, I’m really glad you found the article helpful 🙂 I will pass on your comment to the author. You might also like this recent post on the evolution of cannabis, which covers some of the most recent research on the development of cannabis taxonomy.
With best wishes,
Scarlet
Touche’ ! Ya got it down!