




Die verwirrende Welt der Taxonomie hilft Botanikern, die phylogenetische Reise einer Pflanze zu verstehen. In diesem Artikel untersuchen wir die Phylogenie (evolutionären Beziehungen) von Cannabis und seine Entwicklung in die Familie der Cannabaceae. Wir sprechen über Vorfahren und enge Verwandte von Cannabis und andere Mitglieder der Cannabaceae-Familie.
- Cannabis im kladistischen System
- Die komplexe und verwirrende Welt der Pflanzentaxonomie
- Merkmale der verschiedenen Taxa
- Die Rosidae Superordnung und die Rosaceae Familie
- Die relativ jungen urtikaläischen Rosiden
- Die verwirrende Klassifizierung der Cannabaceae Familie
- Andere erwähnenswerte Cannabaceae-Pflanzen
- Kommen Cannabinoide nur in Cannabis vor?
Die Aufgabe, Milliarden von Pflanzenarten, die seit Beginn des Lebens auf unserem Planeten existieren, zu kategorisieren und katalogisieren, ist eine riesige und äußerst komplexe Aufgabe. Vor der Entwicklung von Gentests war die Klassifizierung auf der Grundlage der wahrgenommenen Ähnlichkeit die einzige Möglichkeit, eine Pflanze ihrem richtigen Taxon zuzuordnen.
Nachdem wir nun die phylogenetische Forschung (evolutionäre Beziehungen zwischen Organismengruppen) entwickelt haben, haben wir eine Grundlage für die Klassifizierung von Pflanzen nach ihren genetischen Ähnlichkeiten – aber das bietet keinen problemlosen Ansatz für eine so komplexe Aufgabe.
Cannabis im kladistischen System
Wir können aber zumindest damit beginnen, die Pflanzen nach einem kladistischen System zu klassifizieren. Das kladistische System beruht auf der Anwesenheit eines einzigen gemeinsamen Vorfahrens. Auf diese Weise enthält jede Klade (Zweig) des phylogenetischen Baumes die Nachkommen eines einzigen gemeinsamen Vorfahren. Wenn viele Pflanzen einen gemeinsamen Vorfahren haben, gelten sie als monophyletisch.
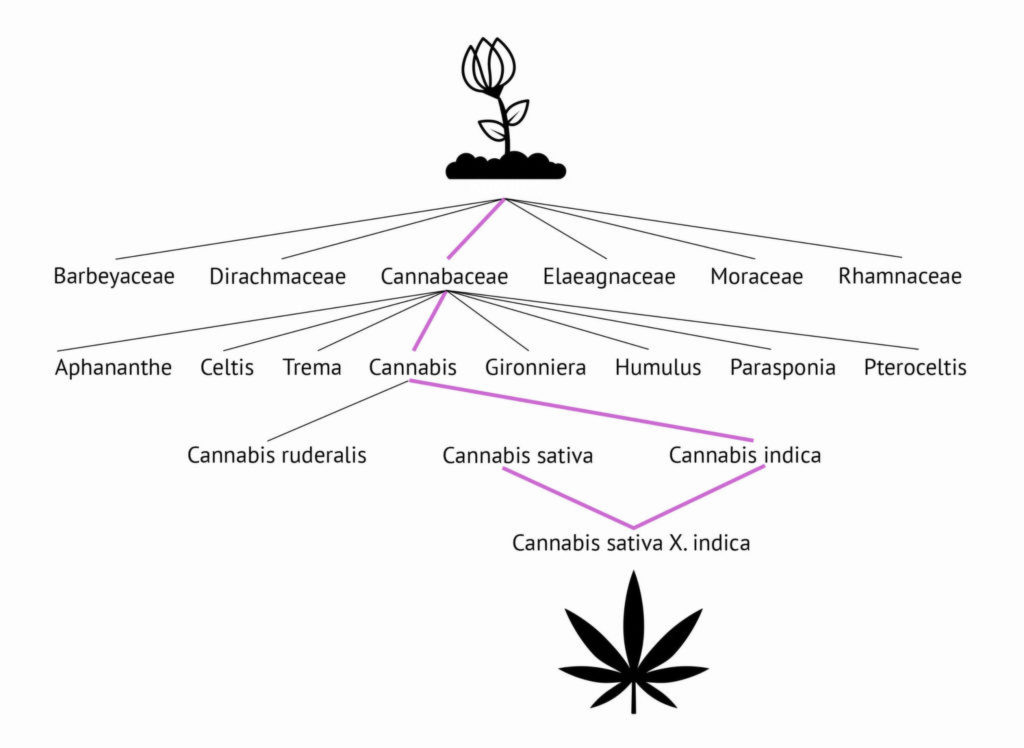
Nach dem Integrierten Taxonomischen Informationssystem (ITIS) gehört die Art Cannabis Sativa L. zur Gattung Cannabis, die zur Familie der Cannabaceae gehört. Die Familie der Cannabaceae bildet zusammen mit drei weiteren Familien die (informelle) Unterordnung der Urticaceen (Brennnesselgewächse), der Ordnung Rosales (Rosenartige). Neben den rosenartigen Brennnesselgewächsen gehören fünf weitere Familien zu den Rosenartigen; dazu gehören die Rosaceae (Rosen) und Rhamnaceae (Sanddorn).
Verwandter Post

Die komplexe und verwirrende Welt der Pflanzentaxonomie
Der Rosenartigen gehört zur informellen Unterklasse (oder Überordnung) Rosanae (oder Rosidae), einer riesigen Kluft, die fast ein Viertel aller blühenden Pflanzen umfasst. Diese „Rosiden“ gehören zur Klasse der Magnoliopsida oder Eudicot, das Mitglied der Infradivision Angiospermae ist. Diese wiederum gehören zur Unterteilung der Spermatophytina, zur Unterteilung der Tracheophyta, zum Infrakreich Streptophyta oder Embryophyta (Landpflanzen), zum Subreich Viridiplantae (grüne Pflanzen).
Das Viridiplantae-Phylum besteht aus Grünalgen sowie allen Landpflanzen. Sie bildet zusammen mit der Phyla von Rhodophyta (Rotalgen) und Glaukophyten (Süßwassermikroalgen) das Archaeplastida-Königreich, das auch als Plantae sensu lato bekannt ist – was „Pflanzen im weiteren Sinne“ bedeutet.
Die Frage, ob Archaeplastida tatsächlich eine höhere Klade ist, die sich aus den Königreichen von Viridiplantae usw. zusammensetzt, ist etwas umstritten.
Merkmale der verschiedenen Taxa
Archaeplastida verfügen alle über Chloroplasten, die zur Energiegewinnung Photosynthese durchlaufen, außerdem haben die Chloroplastenzellen der Archaeplastiden gemeinsame Strukturen, die typischerweise von nur zwei Membranen umgeben sind.
Viridiplantae unterscheidet sich von anderen Phyla in der Archaeplastida-Klasse dadurch, dass sie sowohl Chlorophyll a als auch b produzieren, was ihnen ihre übliche grüne Farbe verleiht. Rhodophyta und Glaukophyten produzieren nur Chlorophyll a.
Embryophyten umfassen alle Landpflanzen und sollen vor etwa 540 Millionen Jahren aus Grünalgen entstanden sein.
Tracheophyta, oder vaskuläre Pflanzen, enthalten Fasergewebe, um Wasser und Mineralien zu leiten, und umfassen Moose und Farne sowie Blütenpflanzen. Spermatophytina sind vaskuläre Pflanzen, die Samen produzieren, und es wird angenommen, dass sie vor etwa 319 Millionen Jahren aus nicht-saatgutführenden Pflanzen entstanden sind.
Angiospermae werden daher aufgrund ihrer gemeinsamen Eigenschaft, Samen zu tragen, die in Schutzstrukturen eingeschlossen sind, klassifiziert; sie alle tragen Blüten, die später die samenhaltigen Früchte produzieren und seit etwa 192 Millionen Jahren auftauchen. Eudikotta sind Angiospermen, die zwei embryonale Blätter exprimieren, im Gegensatz zu Monokotylen, die nur eines produzieren. Eudikotis zeigen außerdem ein charakteristisches dreirilliges Muster auf der Pollenflora.
Verwandter Post

Die Rosidae Superordnung und die Rosaceae Familie
Es wird angenommen, dass die Rosidae vor 108-121 Millionen Jahren erschienen sind und sie sind in der Regel durch ihre auffälligen Blüten erkennbar – die urtikaläischen Rosiden sind bemerkenswerte Ausnahmen von dieser Regel. Forschungen haben gezeigt, dass die jüngsten evolutionären Divergenzen in Richtung der Unauffälligkeit tendiert haben, da die Windbestäubung immer häufiger wurde. Die Rosales-Ordnung umfasst eine große Vielfalt an immergrünen und laubabwerfenden Bäumen, Sträuchern, Reben und Kräutern: So enthält die Familie der Rosaceae eine unglaubliche Vielfalt an wichtigen Arten wie Pfirsich, Apfel, Erdbeere, Mandel und natürlich Rose.

Die relativ jungen urtikaläischen Rosiden
Die urtikaläischen Rosiden, die als relativ junge Ableger des phylogenetischen Baumes gelten, zeichnen sich durch ihre oft unauffälligen und meist unisexuellen Blüten aus (obwohl Ulmenblüten meist bisexuell sind). Neben den Cannabaceae gehören zu den Urticales die Familien Urticaceae (Brennnessel), Ulmaceae (Ulme) und Moraceae (Maulbeere oder Feige). Männliche Blüten sind unauffällig und enthalten keine Blütenblätter, nur Kelchblätter; weibliche Fruchtknoten enthalten eine einzige Eizelle und produzieren einen einzigen Samen.
Urtikaläische Rosiden weisen große Unterschiede in Morphologie und Biogeographie auf und haben bemerkenswerte Anpassungen entwickelt. Die Pflanzen werden in der Regel windbestäubt, obwohl mehrere Mitglieder der Familie der Moraceae von Insekten bestäubt werden. Mehrere Arten innerhalb der Brennnesselfamilie haben die einzigartige Fähigkeit, Pollen explosionsartig in den Wind zu zerstreuen. Früchte können harte, trockene Nüsse sein, wie in Cannabis; fleischige Steinfrüchte, wie bei Traubenkirschen; fleischige Synkonien, einzigartig bei Feigen; oder zusammengesetzte Cluster, wie sie im Maulbeerbaum vorkommen. Interessanterweise wird angenommen, dass mehrere Maulbeerarten milde halluzinogene Eigenschaften aufweisen, und aus der Rinde können hochwertige Fasern gewonnen werden.
Verwandter Post

Die verwirrende Klassifizierung der Cannabaceae Familie
Die etwa 170 Mitglieder der Cannabaceae-Familie haben nur wenige Merkmale, die sie von den Urticales als Ganzes unterscheiden. DNA-Untersuchungen haben gezeigt, dass alle verwandt sind, und sicherlich haben Humulus (Hopfen) und Cannabis offensichtliche Ähnlichkeiten in Bezug auf die Blumenstruktur. Beide enthalten Terpenoide, die für ihren ähnlichen Duft verantwortlich sind, aber Humulus ist eine Weinrebe (Kletterpflanze) und Cannabis ist krautig. Die Gattung Celtis (Traubenkirsche) ist relativ unähnlich: Celtis sind in der Regel hohe Laubbäume mit einfachen Blättern, im Gegensatz zu den zusammengesetzten Palmenblättern bei Humulus und Cannabis. Celtis sind in der Regel nicht zweihäusig und obwohl die Blüten meist unisexuell sind, befinden sie sich auf derselben Pflanze.

Trotz der vielen Unterschiede ähneln die männlichen Blüten vieler Celtis-Arten auffallend denen von Cannabis. Weitere Ähnlichkeiten zwischen den Mitgliedern der Cannabaceae Familie bestehen aus stipule-tragenden Blättern (bei Cannabis sind die Nebenblätter die beiden winzigen Triebe an der Basis jedes Blattes) und Zystolithen oder vergrößerten Blattzellen mit Calciumcarbonatkristallen.
Andere erwähnenswerte Cannabaceae-Pflanzen
Obwohl allgemein angenommen wird, dass Cannabis und Humulus die einzigen beiden Cannabaceae-Gattungen mit kommerziellem Wert sind, gibt es mehrere andere Arten, die in den Regionen, in denen sie heimisch sind, von großem Wert sind.
Verschiedene Celtis-Arten werden entweder für ihr Holz oder als Zierpflanzen angebaut, wie der geschützte südafrikanische Baum C. Africana. Er ist attraktiv, schnellwüchsig und frost- und trockenheitsbeständig. Seine üppigen Blüten und kleinen, süßen Früchte ziehen eine Vielzahl von Vögeln und Insekten in den Garten.
Auch C. sinensis aus Ostasien und C. australis aus Südeuropa werden weitgehend für diese Zwecke angebaut.
Trema orientalis, eine häufige Art, die in den tropischen und warmen gemäßigten Regionen Afrikas, Asiens und Ozeaniens verbreitet ist, wird häufig genutzt. Viele einheimische Populationen bezeichnen ihn als „Holzkohlenbaum“, da er unter anderem für die Holzkohleproduktion verwendet wird. Sein Holz brennt schnell und einfach und ist aufgrund der hohen Zugfestigkeit der Fasern auch für die Herstellung von Papier und Zellstoff geeignet.
Die Fasern der Rinde werden zur Herstellung von Seilen oder Garnen verwendet, und der ausgestoßene Saft kann zur Abdichtung von Angelschnüren verwendet werden. Er wird auch als Tierfutter verwendet. Interessanterweise hat T. orientalis verschiedene traditionelle medizinische Anwendungen, von denen viele denen von Cannabis sehr ähnlich sind – es wird unter anderem zur Behandlung von Ruhr, Entzündungen, Halsschmerzen und Asthma eingesetzt und hat in Versuchen vielversprechende Ergebnisse als Blutzuckerstabilisator bei Diabetes gezeigt.
Aphananthe aspera, eine ostasiatische Art, die auch als Muku-Baum bekannt ist, ist ein hochangesehenes Ornament, das traditionell in klassischen chinesischen Gärten verwendet wird. A. aspera ist auch als Faser- und Holzquelle nützlich, und seine Blätter werden als feines Schleifpapier zum Polieren von Holz und ähnlichen Gegenständen verwendet. Ein besonders feiner, siebenstämmiger A. aspera wird als eines der Naturdenkmäler Südkoreas aufgeführt; dieses sechzehn Meter hohe Exemplar gilt als etwa dreihundert Jahre alt.

Kommen Cannabinoide nur in Cannabis vor?
Die Gattung Cannabis ist offenbar einzigartig innerhalb der Familie der Cannabaceae, da sie Cannabinoide enthält. Es gibt Hinweise darauf, dass Cannabimimetika – cannabinoidähnliche Verbindungen – in anderen Pflanzenarten vorkommen, insbesondere im Sonnenhut, der zur Unterklasse der Asteriden gehört und sich daher seit Millionen von Jahren separat zu Cannabis entwickelt haben muss. Dies kann bedeuten, dass ein Proto-Cannabinoidsystem in einem gemeinsamen Vorfahren beider Pflanzen existierte, bevor die Rosiden- und Asteridenkladen vor etwa 126 Millionen Jahren divergiert sind.
Nach heutigem Kenntnisstand produziert jedoch keine andere Pflanzenart THC oder CBD. Diese Cannabinoide gibt es nur in der Cannabispflanze. Es gibt Unternehmen, die behaupten, CBD aus Quellen wie Limonen aus Zitrusfrüchten und der Rinde von immergrünen Pflanzen aus Südostasien hergestellt zu haben, aber bisher gab es keine unabhängigen, von Experten begutachteten Untersuchungen, um diese Behauptungen zu überprüfen.
Angesichts der großen Unwahrscheinlichkeit, dass eine neue Pflanzenart, die ein voll funktionsfähiges Phytocannabinoidsystem enthält, plötzlich entsteht, scheint die allmähliche Evolution komplexer Cannabinoidsysteme aus rudimentäreren Linien die einzige plausible Erklärung zu sein. Spuren früher Cannabaceae-Arten wurden in kreidehaltigen Fossilien mit einem Alter von bis zu 93,5 Millionen Jahren gefunden, aber es gibt keine konkreten Hinweise darauf, wann die Cannabis-Gattung divergierte.
Die Tatsache, dass keine nahen Verwandten von Cannabis Spuren für ein Cannabinoidsystem zeigen, ist überraschend, aber mit fortschreitender Forschung könnten mehr cannabinoidhaltige Pflanzen entdeckt werden. Dies kann eine weitere Umgestaltung des komplexen und verwirrenden Systems der Klassifizierung von Pflanzen erforderlich machen.

Hallo Danke für diesen Artikel ich hätte aber ne frage und zwar wie die Blüten Formel für eine weibliche cannabis Pflanze aussieht ich lerne gerade Gärtner aber nicht mal meine Lehrer können mir dies beantworten .vielen Dank